
 تويت
تويت
عبور وراثي
Crossing over - Entrecroisement
العبور الوراثي
إن أعداد صبغيات الكائن الحي أقل كثيراً من أعداد مورِّثاته (جيناته) genes التي تحملها، وعلى هذا فإن كل صبغي يمتلك مجموعة من المورثات تدعى مجموعة ارتباطية، وقد أشار إليها ساتونSutton منذ عام 1903. وتتساوى أعداد المجموعات الارتباطية مع أعداد أشفاع الصبغيات، فالإنسان يمتلك 23 شفعاً من الصبغيات والمجموعات الارتباطية، مقابل أربعة أشفاع في ذبابة الخل (الدروزوفيلا ميلانوغاستر Drosophila melanogaster)، وهكذا.
يقصد بالارتباط الوراثي genetic linkageوجود مجموعة من المورثات على الصبغي، ويؤدي ذلك إلى عدم توزعها بشكل مستقل حسب قوانين مندل Mendel [ر. مندل (غريغور ـ)]. وكان مورغنMorgan أول من أثبت الترتيب الخطي للمورثات على الصبغيات، وأشار إلى أن درجة (أو شدة) الارتباط بين المورثات تتوقف على المسافات التي تفصل بينها. أما في حالة الوراثة المندليةMendelian genetics فإن ما يعرف بالتوزع المستقل للصفات يحدث عندما تقع المورثات المسؤولة عنها في صبغيات مختلفة.
العبور الوراثي genetic crossing- over، هو تبادل فيزيائي في مواقع بعض المورثات على الصبغيات القرينة، مؤدياً إلى تكوين اتحادات جديدة (عبورية) تختلف عن الأنماط الأبوية. وتحدث هذه الظاهرة في نسبة معينة من الأعراس الناتجة من أفراد غير متماثلة اللواقح. أما في الحالات العادية، التي لا يحدث فيها العبور الوراثي، فإن المورثات المرتبطة تنتقل كوحدة واحدة من جيل إلى آخر.
من أوائل التجارب التي أوضحت مفهومي الارتباط والعبور، تجربة نفّذها بانِت Punnettوباتسون Bateson على البازلاء، درسا فيها الصفات الآتية:
أزهار حمراء = R-، أزهار بيضاء = rr.
حبوب طلع طويلة = L-، حبوب طلع مستديرة = ll.
ووجدا زيادة في عدد أعراس المورثتين PLوpl بالموازنة مع أعراس المورثتين Pl وpL، وهذا ما توضحه دراسة توزع الصفات في الجيل الثاني وفق الجدول (1).
وتوضح هذه النتائج عدم وجود توزع مستقل للمورثات (ومن ثم الصفات) وحدوث ظاهرتي الارتباط والعبور الوراثيين.
يتضح مما سبق أن الارتباط يمثل علاقة فيزيائية بين المورثات، ومن ثم فإن الحصول على اتحادات جديدة يتم عن طريق انقسام وتبادل فيزيائي بين أشفاع المورثات المتوضعة بترتيب خطي على الصبغيات المتقابلة، إذ تتعلق نسبة العبور بطول المسافة بين المورثتين المتجاورتين، فبازدياد هذه المسافة تزداد فرصة حدوث التصالبchiasma بين شفعي الصبغي، والعكس صحيح. وغالباً ما يكون الارتباط تاماً بين المورثتين اللتين تكونان أقرب ما يمكن من منطقة القُسَيْم (الجزيء) المركزي centromere، علماً بأنه لا يوجد ارتباط تام في معظم الأنواع المتكاثرة جنسياً إلا نادراً.
أشار مورغان عام 1910 إلى أن الحصول على اتحادات جديدة يكون نتيجة تبادل أجزاء بين أشفاع الصبغيات في أثناء الانقسام المنصف (الاختزالي) meiosis، ففي الطور التحضيري الأولprophase I وفي الدور التزاوجي zygotene تقترب الصبغيات المتناظرة ويحصل الاقتران synapsisبين جميع الأجزاء المتشابهة على الصبغيات القرينة ويستمر حتى الدور الضام pachytene، حيث ينقسم كل صبغي إلى كروماتيدتين chromatidsوتصبح الوحدة الثنائية مكونة من أربع كروماتيدات مشكلة ما يسمى الرباعيات tetrads. وقد لوحظ في نهاية هذا الدور أو بداية دور التضاعف diplotene تشكل تصالب واحد أو أكثر على شكل حرف (X)، تتقاطع فيه الكروماتيدات غير الشقيقة معاً، ويلي ذلك حدوث انقسام في منطقة التقاطع، والتحام جديد (الشكل 1).
أ ـ (اليسار): انعزال زوجين من المورثات المحمولة على زوجين من الصبغيات والحصول على نوعين من الأعراس (من دون عبور).
ب ـ (اليمين): الحصول على أربع أنواع مختلفة من الأعراس نتيجة لحدوث عبور مفرد.
تطبيقات العبور الوراثي
رسم الخرائط الصبغية هو من أهم تطبيقات ظاهرة هذا العبور إذ يحدد فيه ترتيب تتابع المورثات في الصبغي الواحد والمسافات الصبغية بينها. وتقدر المسافة الصبغية بين موقعي مورثتين بالمسافة التي تسمح بحدوث نسبة عبور مقدارها 1%. فإذا كانت نسبة العبور 15% بين المورثتين AوB مثلاً، فإن هذا يعني أن المسافة بينهما هي 15وحدة عبورية أو سنتي مورغان Centi Morgan.
ولأن نسبة الاتحادات الجديدة بين المورثتين المرتبطتين ثابتة في ظروف بيئية محددة فإن هذا التكرار الثابت والمسمى بالقيمة العبورية يساعد على رسم الخارطة الصبغية. ويمكن ايضاح طريقة ذلك كما يأتي:
لتحديد مواقع ثلاث مورثات على صبغي معين، تجرى تجربة تدعى تجربة الثلاث نقاط، اعتماداً على العلاقة الهندسية بين ثلاث نقاط واقعة على خط مستقيم، فإذا كانت المورثات AوB وC واقعة على صبغي واحد، فإن المسافة ACهي إما حاصل جمع أو طرح المسافتين AB وBC.
فإذا افترض أن نسبة العبور بين A وB = 5%(أي المسافة بينهما هي خمس وحدات صبغية).
وبين A و C= 7% وبين B وC =12%، أمكن القول إن المورثة A تقع بين B وC على النحو الآتي:
أما إذا كانت نسبة العبور بين B وC مساوية لـ 2% فإن ترتيب هذه المورثات يكون على النحو الآتي:
مثال: الصفات الثلاث «الجسم الأصفر والعيون البيضاء والجناح الملتحم العروق» متنحية ومرتبطة بالجنس في ذبابة الخل، وتوجد بينها قيم العبور الآتية:
1.5% بين مورثتي الجسم الأصفر (y) والعيون البيضاء (w).
5.4% بين مورثتي العيون البيضاء (w) والجناح الملتحم العروق (v).
6.9% بين مورثتي الجسم الأصفر (y) والجناح الملتحم العروق (v).
وعلى ذلك يمكن تعيين مواقع المورثات المذكورة على الصبغي X على النحو الآتي:
وبافتراض أن عبوراً في منطقة صبغية معينة لا علاقة له بعبور في منطقة أخرى من الصبغي ذاته، فإن احتمال حدوث عبورين في وقت واحد في موقعين مختلفين من الصبغي الواحد يكون مساوياً لجداء الاحتمالين المستقلين لكل منهما، وإن حدوث عبور مفرد في منطقة ما من الصبغي يخفض قليلاً أو كثيراً من احتمال حدوث عبور آخر مفرد في منطقة أخرى من الصبغي نفسه وهو ما يعرف بالتداخل (أو التعارض) interference. وعملياً فإن حدوث العبور المزدوج أو المتعدد يخفض كثيراً من نسب العبور.
ساعد تحديد المسافات الوراثية ومن ثم تحديد نسب العبور الحقيقية في رسم الخرائط الصبغية chromosome maps لعدد كبير من الكائنات مثل ذبابة الخل وعدد من البكتريا والذرة الصفراء والقمح والشعير وفول الصويا وغيرها، ومكّن علماء الوراثة والتربية من تحديد المورثات المسؤولة عن الصفات ومعرفة فيما إذا كانت مرتبطة فيما بينها أم لا. ولهذا الموضوع أهمية كبيرة في وضع برامج التحسين الوراثي الهادفة إلى نقل الصفات الاقتصادية سواء عن طريق التهجين أو التطفير أو بوسائل الهندسة الوراثية.
هذا وقد طورت طرائق حديثة وتقانات معقدة لتجديد مواقع المورثات على الصبغيات، واستخدمت بكفاءة عالية في مشروع الجينوم البشري [ر] الذي أعلنت نتائجه في مطلع القرن الجاري.
محمد معلا
Crossing over - Entrecroisement
العبور الوراثي
إن أعداد صبغيات الكائن الحي أقل كثيراً من أعداد مورِّثاته (جيناته) genes التي تحملها، وعلى هذا فإن كل صبغي يمتلك مجموعة من المورثات تدعى مجموعة ارتباطية، وقد أشار إليها ساتونSutton منذ عام 1903. وتتساوى أعداد المجموعات الارتباطية مع أعداد أشفاع الصبغيات، فالإنسان يمتلك 23 شفعاً من الصبغيات والمجموعات الارتباطية، مقابل أربعة أشفاع في ذبابة الخل (الدروزوفيلا ميلانوغاستر Drosophila melanogaster)، وهكذا.
يقصد بالارتباط الوراثي genetic linkageوجود مجموعة من المورثات على الصبغي، ويؤدي ذلك إلى عدم توزعها بشكل مستقل حسب قوانين مندل Mendel [ر. مندل (غريغور ـ)]. وكان مورغنMorgan أول من أثبت الترتيب الخطي للمورثات على الصبغيات، وأشار إلى أن درجة (أو شدة) الارتباط بين المورثات تتوقف على المسافات التي تفصل بينها. أما في حالة الوراثة المندليةMendelian genetics فإن ما يعرف بالتوزع المستقل للصفات يحدث عندما تقع المورثات المسؤولة عنها في صبغيات مختلفة.
العبور الوراثي genetic crossing- over، هو تبادل فيزيائي في مواقع بعض المورثات على الصبغيات القرينة، مؤدياً إلى تكوين اتحادات جديدة (عبورية) تختلف عن الأنماط الأبوية. وتحدث هذه الظاهرة في نسبة معينة من الأعراس الناتجة من أفراد غير متماثلة اللواقح. أما في الحالات العادية، التي لا يحدث فيها العبور الوراثي، فإن المورثات المرتبطة تنتقل كوحدة واحدة من جيل إلى آخر.
من أوائل التجارب التي أوضحت مفهومي الارتباط والعبور، تجربة نفّذها بانِت Punnettوباتسون Bateson على البازلاء، درسا فيها الصفات الآتية:
أزهار حمراء = R-، أزهار بيضاء = rr.
حبوب طلع طويلة = L-، حبوب طلع مستديرة = ll.
|
|||||||||||||||||||||||||||||||||||
| الجدول (1) |
وتوضح هذه النتائج عدم وجود توزع مستقل للمورثات (ومن ثم الصفات) وحدوث ظاهرتي الارتباط والعبور الوراثيين.
يتضح مما سبق أن الارتباط يمثل علاقة فيزيائية بين المورثات، ومن ثم فإن الحصول على اتحادات جديدة يتم عن طريق انقسام وتبادل فيزيائي بين أشفاع المورثات المتوضعة بترتيب خطي على الصبغيات المتقابلة، إذ تتعلق نسبة العبور بطول المسافة بين المورثتين المتجاورتين، فبازدياد هذه المسافة تزداد فرصة حدوث التصالبchiasma بين شفعي الصبغي، والعكس صحيح. وغالباً ما يكون الارتباط تاماً بين المورثتين اللتين تكونان أقرب ما يمكن من منطقة القُسَيْم (الجزيء) المركزي centromere، علماً بأنه لا يوجد ارتباط تام في معظم الأنواع المتكاثرة جنسياً إلا نادراً.
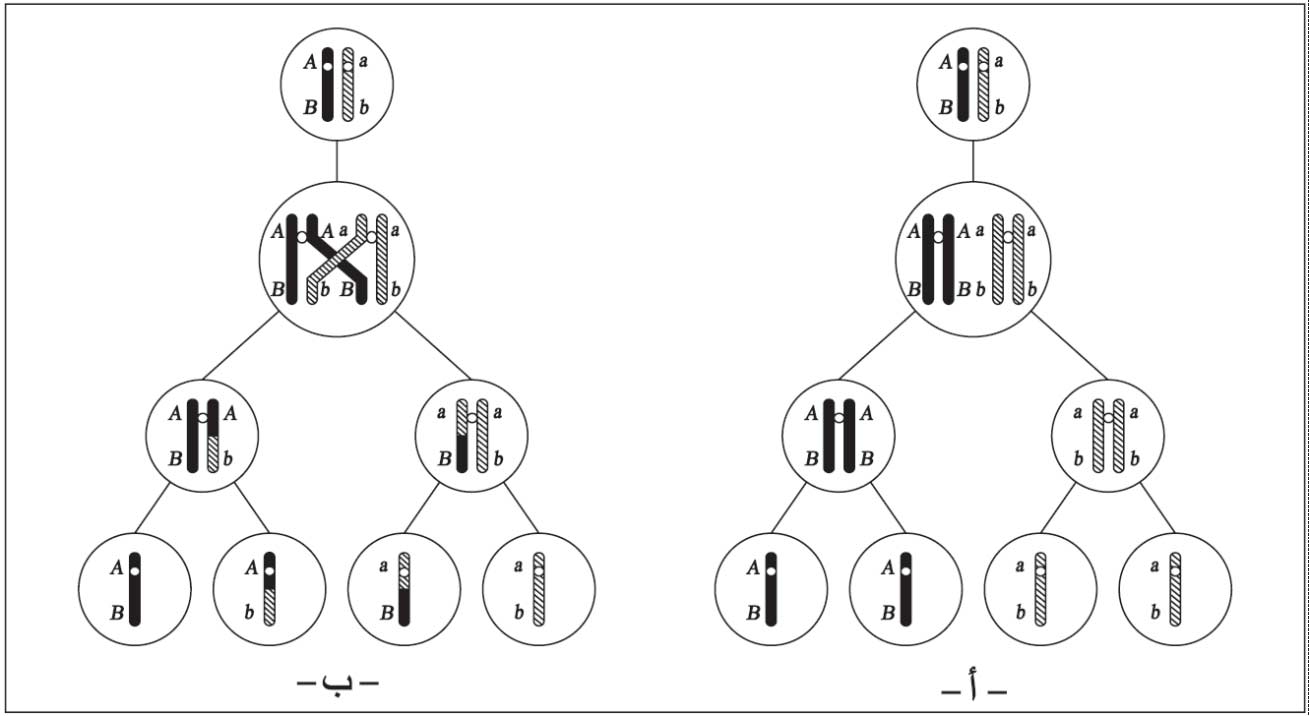 |
| الشكل (1) |
أ ـ (اليسار): انعزال زوجين من المورثات المحمولة على زوجين من الصبغيات والحصول على نوعين من الأعراس (من دون عبور).
ب ـ (اليمين): الحصول على أربع أنواع مختلفة من الأعراس نتيجة لحدوث عبور مفرد.
تطبيقات العبور الوراثي
رسم الخرائط الصبغية هو من أهم تطبيقات ظاهرة هذا العبور إذ يحدد فيه ترتيب تتابع المورثات في الصبغي الواحد والمسافات الصبغية بينها. وتقدر المسافة الصبغية بين موقعي مورثتين بالمسافة التي تسمح بحدوث نسبة عبور مقدارها 1%. فإذا كانت نسبة العبور 15% بين المورثتين AوB مثلاً، فإن هذا يعني أن المسافة بينهما هي 15وحدة عبورية أو سنتي مورغان Centi Morgan.
ولأن نسبة الاتحادات الجديدة بين المورثتين المرتبطتين ثابتة في ظروف بيئية محددة فإن هذا التكرار الثابت والمسمى بالقيمة العبورية يساعد على رسم الخارطة الصبغية. ويمكن ايضاح طريقة ذلك كما يأتي:
لتحديد مواقع ثلاث مورثات على صبغي معين، تجرى تجربة تدعى تجربة الثلاث نقاط، اعتماداً على العلاقة الهندسية بين ثلاث نقاط واقعة على خط مستقيم، فإذا كانت المورثات AوB وC واقعة على صبغي واحد، فإن المسافة ACهي إما حاصل جمع أو طرح المسافتين AB وBC.
فإذا افترض أن نسبة العبور بين A وB = 5%(أي المسافة بينهما هي خمس وحدات صبغية).
وبين A و C= 7% وبين B وC =12%، أمكن القول إن المورثة A تقع بين B وC على النحو الآتي:
 |
 |
1.5% بين مورثتي الجسم الأصفر (y) والعيون البيضاء (w).
5.4% بين مورثتي العيون البيضاء (w) والجناح الملتحم العروق (v).
6.9% بين مورثتي الجسم الأصفر (y) والجناح الملتحم العروق (v).
وعلى ذلك يمكن تعيين مواقع المورثات المذكورة على الصبغي X على النحو الآتي:
 |
ساعد تحديد المسافات الوراثية ومن ثم تحديد نسب العبور الحقيقية في رسم الخرائط الصبغية chromosome maps لعدد كبير من الكائنات مثل ذبابة الخل وعدد من البكتريا والذرة الصفراء والقمح والشعير وفول الصويا وغيرها، ومكّن علماء الوراثة والتربية من تحديد المورثات المسؤولة عن الصفات ومعرفة فيما إذا كانت مرتبطة فيما بينها أم لا. ولهذا الموضوع أهمية كبيرة في وضع برامج التحسين الوراثي الهادفة إلى نقل الصفات الاقتصادية سواء عن طريق التهجين أو التطفير أو بوسائل الهندسة الوراثية.
هذا وقد طورت طرائق حديثة وتقانات معقدة لتجديد مواقع المورثات على الصبغيات، واستخدمت بكفاءة عالية في مشروع الجينوم البشري [ر] الذي أعلنت نتائجه في مطلع القرن الجاري.
محمد معلا